forming a star-like chitinous centre in section, each lateral eye of Scorpio has several rhabdoms of five or less rhabdomeres, indicating that the Limulus lateral eye-unit is more specialized than the detached lateral eyelet of Scorpio, so as to present a coincidence of one lens with one rhabdom. Numerous rhabdomeres (grouped as rhabdoms in Limulus) are found in the retinal layer of the central eyes also.

|
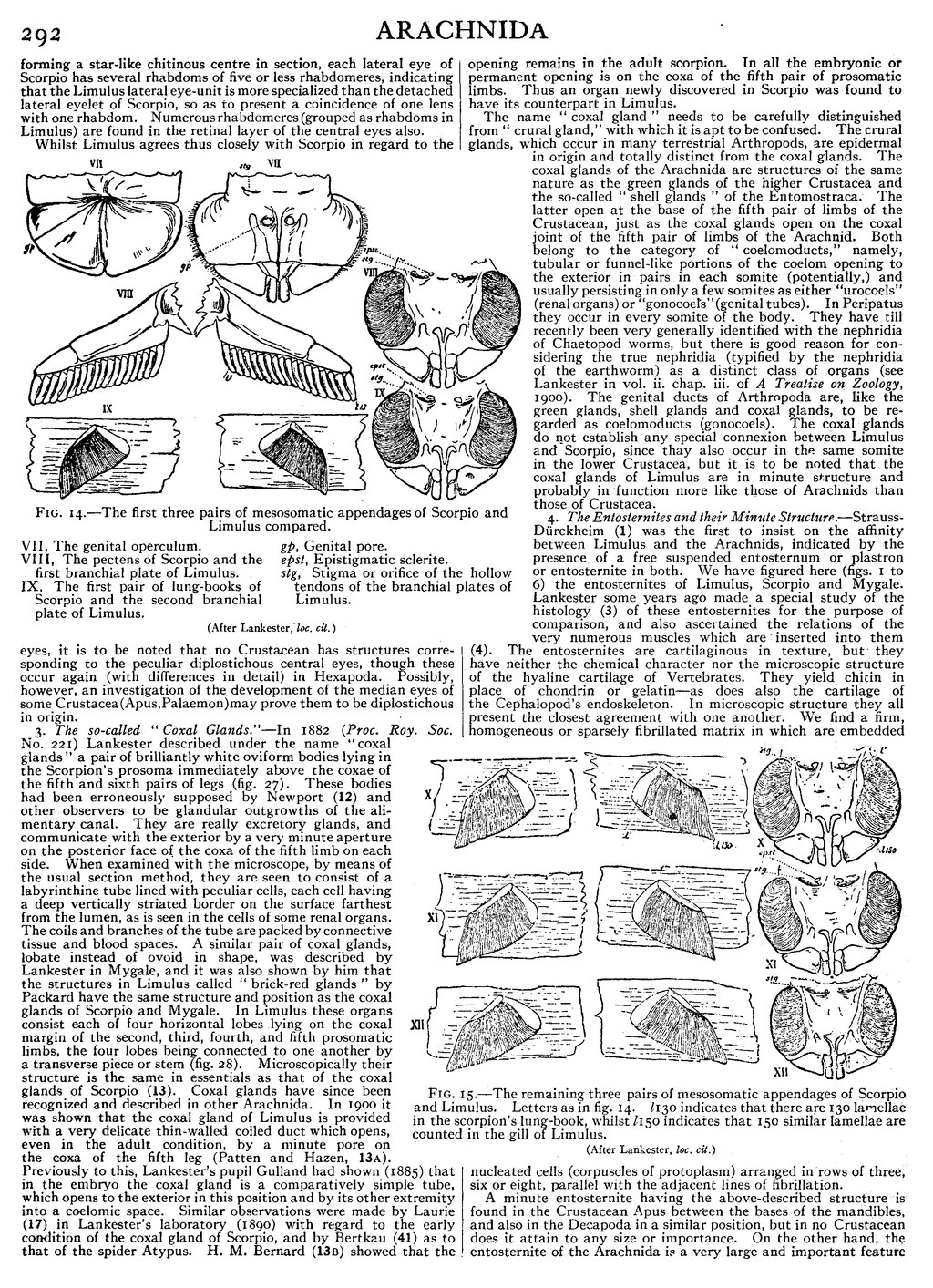
Fig. 14.—The first three pairs of mesosomatic appendages of Scorpio and Limulus compared. |
| VII, The genital operculum. | gp, Genital pore. |
| VIII, The pectens of Scorpio and the first | epst, Epistigmatic sclerite. |
| branchial plate of Limulus. | stg, Stigma or orifice of the hollow tendons of the |
| IX, The first pair of lung-books of Scorpio and the | branchial plates of Limulus. |
| second branchial plate of Limulus. |
(After Lankester, loc. cit.)
Whilst Limulus agrees thus closely with Scorpio in regard to the eyes, it is to be noted that no Crustacean has structures corresponding to the peculiar diplostichous central eyes, though these occur again (with differences in detail) in Hexapoda. Possibly, however, an investigation of the development of the median eyes of some Crustacea (Apus, Palaemon) may prove them to be diplostichous in origin.
3. The so-called “Coxal Glands.”—In 1882 (Proc. Roy. Soc. No. 221) Lankester described under the name “coxal glands” a pair of brilliantly white oviform bodies lying in the Scorpion’s prosoma immediately above the coxae of the fifth and sixth pairs of legs (fig. 27). These bodies had been erroneously supposed by Newport (12) and other observers to be glandular outgrowths of the alimentary canal. They are really excretory glands, and communicate with the exterior by a very minute aperture on the posterior face of the coxa of the fifth limb on each side. When examined with the microscope, by means of the usual section method, they are seen to consist of a labyrinthine tube lined with peculiar cells, each cell having a deep vertically striated border on the surface farthest from the lumen, as is seen in the cells of some renal organs. The coils and branches of the tube are packed by connective tissue and blood spaces. A similar pair of coxal glands, lobate instead of ovoid in shape, was described by Lankester in Mygale, and it was also shown by him that the structures in Limulus called “brick-red glands” by Packard have the same structure and position as the coxal glands of Scorpio and Mygale. In Limulus these organs consist each of four horizontal lobes lying on the coxal margin of the second, third, fourth, and fifth prosomatic limbs, the four lobes being connected to one another by a transverse piece or stem (fig. 28). Microscopically their structure is the same in essentials as that of the coxal glands of Scorpio (13). Coxal glands have since been recognized and described in other Arachnida. In 1900 it was shown that the coxal gland of Limulus is provided with a very delicate thin-walled coiled duct which opens, even in the adult condition, by a minute pore on the coxa of the fifth leg (Patten and Hazen, 13a). Previously to this, Lankester’s pupil Gulland had shown (1885) that in the embryo the coxal gland is a comparatively simple tube, which opens to the exterior in this position and by its other extremity into a coelomic space. Similar observations were made by Laurie (17) in Lankester’s laboratory (1890) with regard to the early condition of the coxal gland of Scorpio, and by Bertkau (41) as to that of the spider Atypus. H. M. Bernard (13b) showed that the opening remains in the adult scorpion. In all the embryonic or permanent opening is on the coxa of the fifth pair of prosomatic limbs. Thus an organ newly discovered in Scorpio was found to have its counterpart in Limulus.
The name “coxal gland” needs to be carefully distinguished from “crural gland,” with which it is apt to be confused. The crural glands, which occur in many terrestrial Arthropods, are epidermal in origin and totally distinct from the coxal glands. The coxal glands of the Arachnida are structures of the same nature as the green glands of the higher Crustacea and the so-called “shell glands” of the Entomostraca. The latter open at the base of the fifth pair of limbs of the Crustacean, just as the coxal glands open on the coxal joint of the fifth pair of limbs of the Arachnid. Both belong to the category of “coelomoducts,” namely, tubular or funnel-like portions of the coelom opening to the exterior in pairs in each somite (potentially,) and usually persisting in only a few somites as either “urocoels” (renal organs) or “gonocoels” (genital tubes). In Peripatus they occur in every somite of the body. They have till recently been very generally identified with the nephridia of Chaetopod worms, but there is good reason for considering the true nephridia (typified by the nephridia of the earthworm) as a distinct class of organs (see Lankester in vol. ii. chap. iii. of A Treatise on Zoology, 1900). The genital ducts of Arthropoda are, like the green glands, shell glands and coxal glands, to be regarded as coelomoducts (gonocoels). The coxal glands do not establish any special connexion between Limulus and Scorpio, since they also occur in the same somite in the lower Crustacea, but it is to be noted that the coxal glands of Limulus are in minute structure and probably in function more like those of Arachnids than those of Crustacea.

|
| Fig. 15.—The remaining three pairs of mesosomatic appendages of Scorpio and Limulus. Letters as in fig. 14. l130 indicates that there are 130 lamellae in the scorpion’s lung-book, whilst l150 indicates that 150 similar lamellae are counted in the gill of Limulus. (After Lankester, loc. cit.) |
4. The Entosternites and their Minute Structure.—Strauss-Dürckheim (1) was the first to insist on the affinity between Limulus and the Arachnids, indicated by the presence of a free suspended entosternum or plastron or entosternite in both. We have figured here (figs. 1 to 6) the entosternites of Limulus, Scorpio and Mygale. Lankester some years ago made a special study of the histology (3) of these entosternites for the purpose of comparison, and also ascertained the relations of the very numerous muscles which are inserted into them (4). The entosternites are cartilaginous in texture, but they have neither the chemical character nor the microscopic structure of the hyaline cartilage of Vertebrates. They yield chitin in place of chondrin or gelatin—as does also the cartilage of the Cephalopod’s endoskeleton. In microscopic structure they all present the closest agreement with one another. We find a firm, homogeneous or sparsely fibrillated matrix in which are embedded nucleated cells (corpuscles of protoplasm) arranged in rows of three, six or eight, parallel with the adjacent lines of fibrillation.

|
|  |
| Fig. 16.—Diagram to show the way in which an outgrowing gill-process bearing blood-holding lamellae, may give rise, if the sternal body wall sinks inwards, to a lung-chamber with air-holding lamellae.I is the embryonic condition. bs, Blood sinus. L is the condition of outgrowth with gl, gill lamellae. A is the condition of in-sinking of the sternal surface and consequent enclosure of the lamelligerous surface of the appendage in a chamber with narrow orifice—the pulmonary air-holding chamber. pl, Pulmonary lamellae.bs, Blood sinus. (After Kingsley.) |
Fig. 17.—Embryo of scorpion, ventral view showing somites and appendages.sgc, Frontal groove. sa, Rudiment of lateral eyes. obl, Camerostome (upper lip). so, Sense-organ of Patten. PrGabp1, Rudiment of the appendage of the praegenital somite which disappears. abp2, Rudiment of the right half of the genital operculum. abp3, Rudiment of the right pecten. abp4 to abp7. Rudiments of the four appendages which carry the pulmonary lamellae. I to VI, Rudiments of the six limbs of the prosoma. VIIPrG, The evanescent praegenital somite. VIII, The first mesosomatic somite or genital somite. IX, The second mesosomatic somite or pectiniferous somite. X to XIII, The four pulmoniferous somites. XIV, The first metasomatic somite. (After Brauer, Zeitsch. wiss. Zool., vol. lix., 1895.) |
{kind=link}